Introduction
Hearing loss (HL) is a common sensorineural disorder, with an incidence of 1 in every 500-1,000 children [1]. At least half of the HL cases can be attributed to genetic factors and more than two-third of these individuals are classified as autosomal recessive non-syndromic hearing loss (ARNSHL) [2]. Non-syndromic or syndromic HL may be caused by environmental factors in configuration with a certain genotype. But it should be noted that the environmental factors are almost ineffective on the gap junction protein beta 2 (GJB2) gene. Because HL is extremely heterogeneous, studying large-sized families from different ethnicities such as the Middle East populations are helpful [3].
At least 42 genetic loci are associated with syndromic forms and over 163 genetic loci are associated with non-syndromic HL (http://hereditaryhearingloss.org/). The inheritance of the disease may be autosomal recessive, autosomal dominant, X-linked recessive and mitochondrial [4]. Based on different investigations, mutations of GJB2 gene are the most common cause of ARNSHL worldwide (e.g. Iran).
The connexin 26 (Cx26) gap junctional proteins are encoded by GJB2 gene. Gap junctions are clusters of intercellular channels for cell-to-cell diffusion of small molecules [5]. These proteins exist in different organs and tissues. For example, the second messenger inositol 1,4,5-trisphosphate (IP3) are in cochlea which transmit signals to intracellular compartments and are essential in perceiving sounds, originally assigned to chromosome 13q12. It accounts for about 50% of this common type of HL [6].
The c.35delG mutation that has been tested in recombinant expression systems confirmed loss of function due to altered sorting, also this mutation, is the most frequent recessive Cx26 mutation in Caucasoid families, which results in a frameshift and premature termination of the protein. The pattern of GJB2 allele distribution has a considerable diversity among different races and ethnic groups. A variant-specific, the c.35delG mutation, accounts for >70% of the disease alleles in the northern and southern Europe as well as American Caucasian, carrier frequencies range from 1.3% to 2.8% [7].
However, the high spectrum and prevalence of GJB2 mutations in exon 2 (coding) exists but the prevalence of this mutations are also significantly evident in exon 1 (non-coding) [8]. The c.-23+1G>A and c.299-300delAT mutations were already detected respectively in exon 1 and exon 2 of GJB2, the c.-23+1G>A is a splice site mutation [9], and the c.299-300delAT mutation was reported to be the severe phenotype of ARNSHL in this population.
The c.299-300delAT mutation caused that gap junction proteins cannot form in the plasma membrane. These disabled proteins were remained in the endoplasmic reticulum (ER), it proposed that ER stress (ERS) and followed of ERS-induced cell death may be responsible for HL [10].
The c.299-300delAT mutation is a frameshift that caused an altered amino acid sequence. Studies showed that c.-23+1G>A produced non-functional mRNA [11]. The c.-23+1G>A mutation was shown in different frequencies between various populations including Mongolian, Yakut, Czech, Hungarian, Turkish and Iranian. However, they do not provide a sufficient data on the frequency of GJB2 mutations with genotype-phenotype correlation, in certain ethnic groups such as Kurds [12]. This study focuses on assessing the contribution of GJB2 mutations (exons 1 and 2) for developing an early onset of HL. Moreover, it aims to determine the mutation profile with genotype-phenotype correlation in hearing-impaired Kurdish participants.
Subjects and Methods
Subjects
In this study 45 Kurdish families with the ARNSHL were selected from a genetic counseling clinic in Iran. ARNSHL individuals were selected by screening the family pedigrees, the patients’ audiograms, the medical records, and also the individual information of each patient in these families. They were, therefore, 45 non-related families. All the family members were interviewed after signing a written consent form. The criteria for choosing individuals to take part in the study were as follows: 1) Autosomal recessive pattern of inheritance should be suspected; 2) Clinical and demographic information should be provided; 3) There must be no syndromic (signs and symptoms other than HL) or environmental evidence (infections such as rubella, meningitis, and ototoxic drugs); and 4) At least two patients should exist in the family.
The Review Board of Shahrekord University of Medical Sciences, Shahrekord, Iran, confirmed this study in 2016 (IR.SKUMS.REC.1395.105). After filling out an informed consent form, about 5 mL of peripheral blood was collected in tubes containing EDTA (0.5 M) from all ARNSHL family members. Also in this study, we collected samples of three generations in each family (grandparents and cousins were included).
DNA extraction
DNA was extracted using a DNA extraction kit (DNPTM, CinnaGen, Tehran, Iran). The quality and quantity of DNA samples can be checked with the Nanodrop spectrophotometer (Thermo Scientific NanoDrop 1000 Spetrophotometer, Thermo Scienific, Wilmington, NC, USA).
Genetic analysis of GJB2
Two methods of mutation detection including genetic linkage analysis followed by DNA sequencing of GJB2 were used. Altogether 45 families participated in parallel in this study.
Amplification of the short tandem repeat markers by linkage analysis
PCR reaction of three primer pairs for three DFNB1 short tandem repeat (STR) markers was performed (Table 1). The amplified conditions were as follows: reaction volume was 15 μL with a final concentration of: ddH2O 7.5 μL, master mix 5.5 μL, Primer F 0.5 μL (20 pmol/μL), Primer R 0.5 μL (20 pmol/μL), and template genomic DNA 1 μL (100 ng/μL).
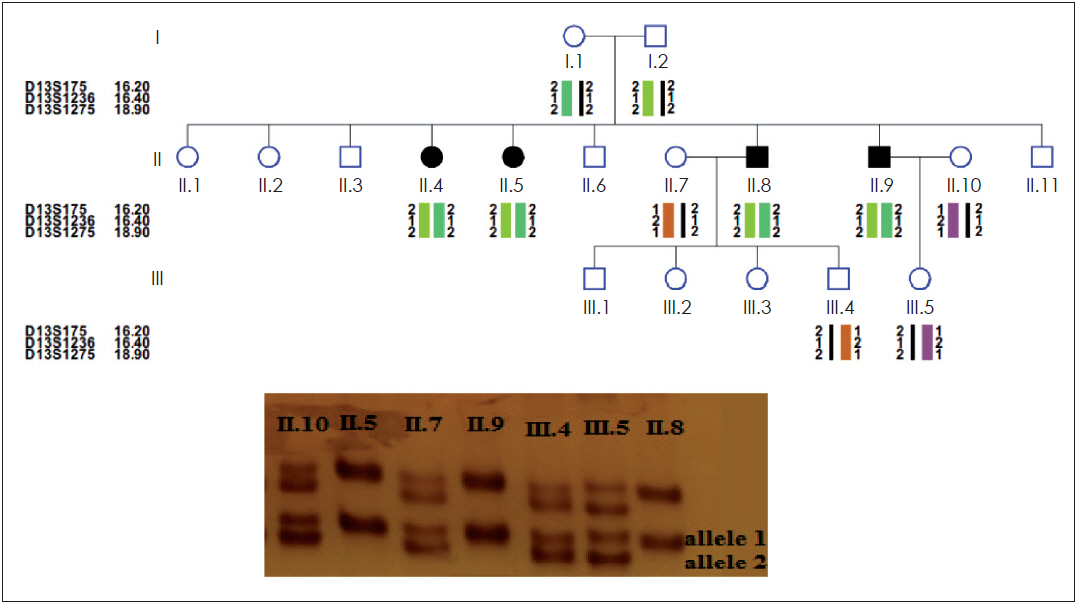
The PCR-reactions were performed in a thermal cycler (Program Temp Control System PC-700, ASTEC, Fukuoka, Japan) with the following conditions: 95°C for 3 minutes, 30 cycles including 95°C for 30 seconds, 57°C for 30 seconds, 72°C for 30 seconds, and finally 72°C for 6 minutes. PCR amplicons were analyzed on a 15% polyacrylamide gel electrophoresis [13] (Fig. 1). Haplotype and LOD score analysis were used to infer genetic linkage to DFNB1 as previously described (Fig. 1).
The 9 families linked to DFNB1 locus were further analyzed for mutation detection but GJB2 was sequenced in each of the tests performed on 45 families to confirm linkage analysis (to carry out a large project and to investigate other effective genes in HL by linkage analysis). The mutations were detected in the 9 families linked and so our results confirm the accuracy of linkage analysis.
Mutation analysis of GJB2 gene in patients by sequence analysis
The GJB2 gene sequencing was performed on one patient in each of the families. Genomic DNA was PCR amplified for sequencing using the following primer pairs: F2: 5' CTC CCT GTT CTG TCC TAG CT 3', R2: 5' CTC ATC CCT CTC ATG CTG TC 3' covering the exon 2 of GJB2. To amplify the exon 1 of GJB2 gene, the following primers were used: F1: 5' CGT CTT TGG GGG TGT TGC TT 3', R1: 5' CAT GAA GAG GGC GTA CAA GTT AGA A 3'.
The amplification conditions were as follows: reaction volume was 25 μL with a final concentration of: ddH2O 18 μL, MgCl2 1.7 μL (100 mM), dNTPs 0.5 μL (10 mM), PCR buffer 2.5 μL, Taq polymers 0.3 μL (3 units/μL), primer F 0.5 μL (20 pmol/μL), primer R 0.5 μL (20 pmol/μL), and template genomic DNA 1 μL (100 ng/μL). The PCR-reaction was performed in a thermal cycler (Program Temp Control System PC-700, ASTEC) at 95°C for 4 minutes, 35 cycles including 95°C for 30 seconds, 60°C for 30 seconds, 72°C for 24 seconds, and finally 72°C for 5 minutes. Sequencing was carried out in SEQLAB (Sequence Laboratories, Gottingen, Germany).
The audiological study of GJB2 mutation positive patients
For each patient (3 to 35 years old), a complete database of the patient’s medical conditions was reviewed to understand the cause of HL, no environmental factors such as maternal to fetal infections, prenatal complications, bacterial or viral infection (meningitis), prenatal and postnatal ototoxic drugs such as aminoglycosides were involved in HL. All the affected family members underwent an ear examination and a general health check to determine the type of HL.
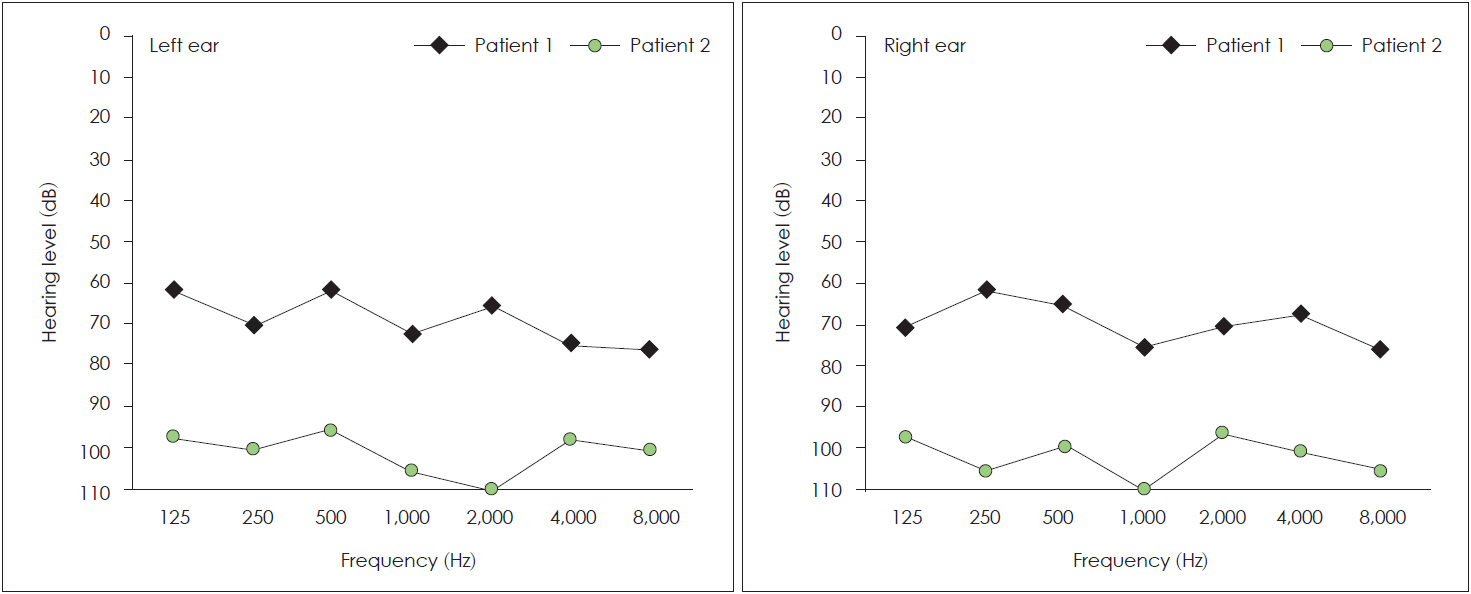
Individuals were also screened for vision assessment, proteinuria, and haematuria. Hearing sensitivity was measured by pure-tone audiometric test (PTA) for air and bone conduction at different frequencies (250-8,000 Hz) in all patients. The degree of HL was classified according to the PTA average at 500, 1,000, 2,000 and 4,000 Hz: mild ≥21 dB, moderate ≥46 dB, moderately severe ≥61 dB, severe ≥76 dB and profound ≥97 dB [14].
Results
In this research 45 Kurdish families were selected to determine the genetic and molecular basis of hearing impairment. HL state was autosomal recessive in this research. HL occurrences may vary in range from mild to profound. Linkage analysis was performed with 3 STR markers (Table 1). Linkage analysis and sequencing were performed for all 45 families who participated in the study but only 9 families were linked to the locus DFNB1 and their mutations were detected by sequencing analysis. So our results confirm the accuracy of linkage analysis.
The c.35delG mutation was detected in 17.7% (8/45) of Kurdish population and the allele frequency was 13.33% (12 positives in 90 alleles). In the 45 families analyzed, the homozygous state was observed for this mutation in 8.8% of the Kurdish population, the heterozygous state was observed in 8.8% and 82.4% of these individual have no c.35delG mutation.
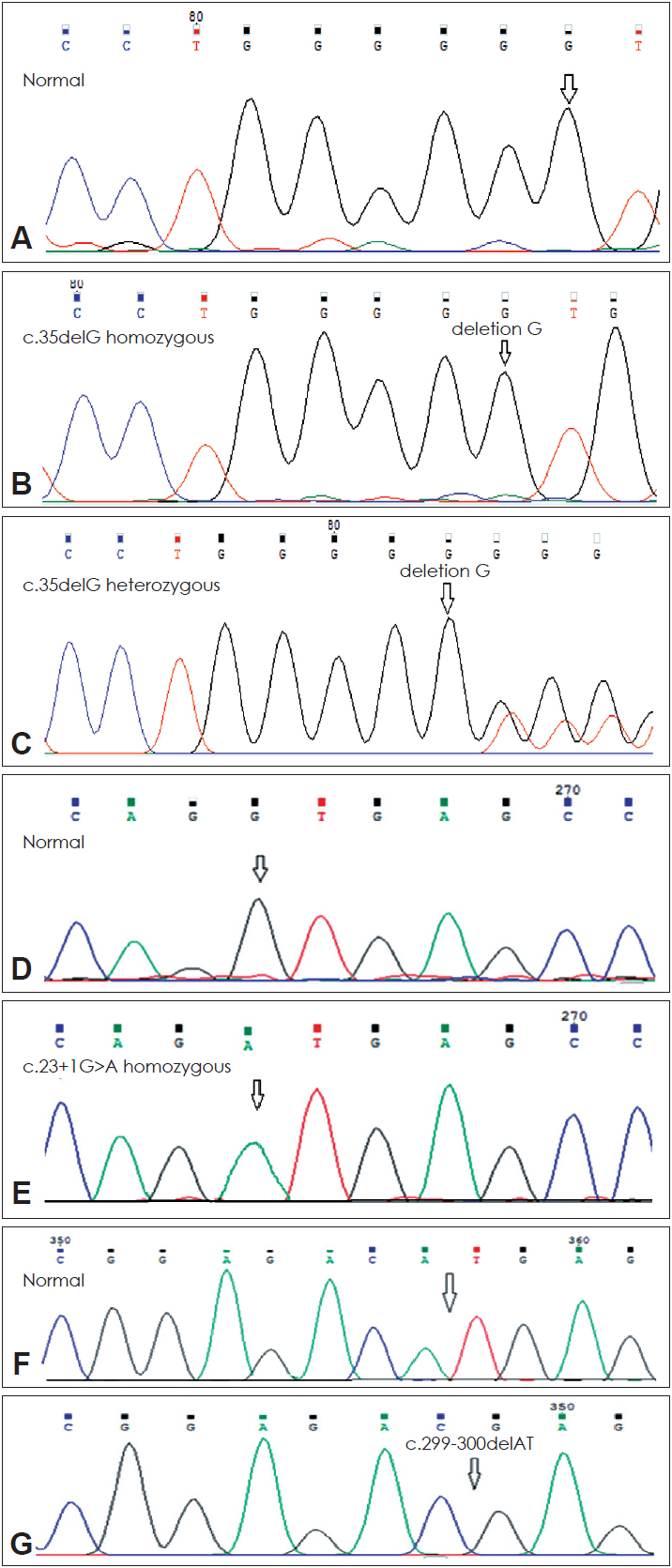
Furthermore, in the next step of the study all individuals who were not c.35delG positive and 4 heterozygous patients for this mutation were analyzed to detect other mutations in the entire coding and exon 1 (noncoding) of GJB2 gene using direct genomic DNA sequencing (Fig. 2). In 8.8% (4/45) of the families, homozygosity for the c.23+1G>A mutation was found in exon 1, also these 4 families had heterozygosity for the c.35delG mutation (Table 2). In addition, 2.2% (1/45) of the families had homozygosity for the c.299-300delAT mutation in exon 2 (Fig. 2). This is the first reported mutation in Kurdish population.
Genotype-phenotype correlation of GJB2 patients
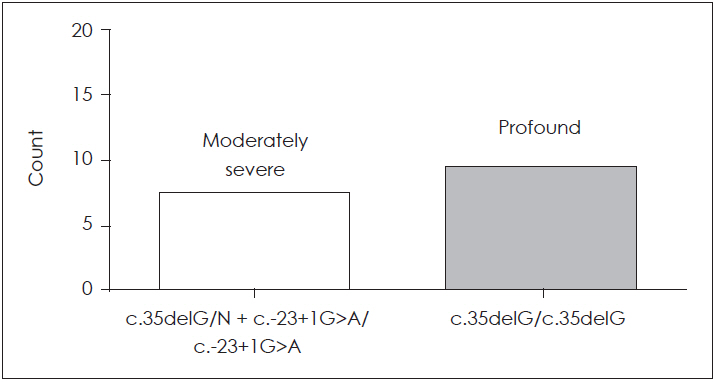
Patients with (c.35delG/N)+(c.-23+1G>A/c.-23+1G>A) genotype and c.35delG homozygous genotype were investigated audiologically. This analysis was carried out on 19 patients from 8 families with GJB2 mutations. Ten patients from these families with profound HL (≥97 dB) at 500, 1,000, 2,000 and 4,000 Hz had c.35delG homozygous mutation (4 families), and 9 other patients (4 families) with moderately severe HL (=61 dB) (Fig. 3) had (c.35delG/N)+(c.-23+1G>A/c.-23+1G>A) genotype (p<0.001) (Fig. 4). Fisher’s exact test was used to detect a positive genotype-phenotype correlation in two genotypes (Table 3). A patient with severe phenotype (≥76 dB) had the c.299-300delAT homozygous genotype.
Discussion
In the present research, the contribution of GJB2 mutations was associated to the HL in nine out of the 45 ARNSHL Kurdish families. We developed a simple, accurate method to determine the c.35delG mutation. Three GJB2 mutations were determined in the present research. The c.35delG mutation, c.-23+1G>A mutation, and c.299-300delAT mutation (first report in Kurdish population) have already been investigated [11]. GJB2 mutations were detected in 9 ARNSHL (20%) patients which was similar to the results of the previous investigations in other regions of Iran [4,15].
The severity of the HL in these 9 patients varied from moderately severe to profound. This study determined the correlation between the phenotype and the GJB2 mutations. Patients with c.35delG/N mutation may have a homozygous different pathogenic mutation in other places. Then after sequencing of the entire coding and noncoding sequence of GJB2 gene, homozygous c.299-300delAT (2.2%) and c.-23+1G>A (8.8%) mutations were observed. Based on these findings, the ARNSHL phenotype in families with c.35delG heterozygote state associated to c.-23+1G>A homozygous state was moderately severe, while families with homozygous c.35delG (8.8%) mutation had profound HL.
Our results detected the contribution of common GJB2 mutations in Kurdish population. Various investigations had determined that the c.35delG mutation is the most common in many ethnic groups, this mutation accounts for 85% of the mutations in GJB2 gene [6]. It is probable that the c.35delG mutation originated in ancient Greece and was subsequently propagated in the Mediterranean [16]. The c.35delG is the most common mutation in the Mediterranean [17], which shows genetic heterogeneity of HL.
The frequency of the c.35delG mutation in the Oman deaf population is null [18], as in African Americans [19], Ghanaian [20], and Kenyan populations [21]. The frequency of this mutation is less than 4% in Australia [22] and Sudan (2.7%); in Iran it is similar to other Mediterranean populations. Also, the main mutations in the GJB2 gene in Dagestan were represented by three forms typical for West Asia: c.35delG mutation (22% of all mutant alleles), deltaE120 mutation (22%), and c.-23+1G>A (44%) were found in a compound heterozygous or a heterozygous state [23], and in Çukurova region of turkey the c.35delG mutation was known as the most frequent mutation of GJB2 gene and had the most frequently mutation (5.5%) in patients with HL [24].
Variation in frequency between different ethnic groups can be the result of human migrations; the high prevalence of the c.35delG mutation in Caucasian populations is possibly the result of a founder effect rather than a mutational hot spot [25]. The Iranian population is composed of various ethnic groups; the prevalence of this mutation differs between Iranian population with a higher slope of mutation frequency from south-tonorth (0% to 33%), and from east-to-west (8% to 8.8%) [12,15].
As a result, GJB2 gene related HL has a low portion of ARNSHL in our population comparing to other ones [26]. The high percentage in northwest and center of Iran may be due to immigration of people in some cities in Iran or may be due to the consanguinity or ascertainment among different ethnic groups. The observations support the presence of a founder effect in northern Iran (compared to 43% in Anatolia) [19]. Based on the population frequencies of the c.35delG mutation and tribes migrating from central Asia to Anatolia 10 centuries ago, we think that the c.35delG mutation probably arose in northwest of Iran [19].
The c.-23+1G>A and c.299-300delAT mutations are among the top ten GJB2 mutations responsible for HL within various populations [14]. Functional studies of c.-23+1G>A mutation have not obtained detectable GJB2 protein and mRNA [11]. Homozygous c.-23+1G>A mutation reported in Iran’s ethnic groups have a different percentage (2.5% average), in another study [27], which is lower than our observation (8.8%).
The (c.35delG/N)+(c.-23+1G>A/c.-23+1G>A) genotype was less severe than that of c.35delG homozygous (p<0.001). We conclude that c.-23+1G>A homozygous mutation is often restricted in the Kurdish population; that shows a founder effect for this mutation. Also, we could identify c.299-300delAT homozygous mutation in one Kurdish family for the first time in Iran who had severe phenotype ARNSHL. The c.299-300delAT mutation is a frameshift that caused an altered amino acid sequence from codon 100, leading to a stop at codon 113 (Fig. 5).
This mutation was first reported as a heterozygote variant related to ARNSHL [28]. Another study found this mutation in homozygote form [29], and then reported it in Turkey [30], and Iran [12]. These findings suggested the involvement of other genetic or environmental causes playing an important role in manifesting any disorders in this population. So far, etiology of this disorder remains largely unknown; the presence of modifier gene or other connexin genes can be involved in HL.
Consequently, more investigations should be conducted to search the existence of pathogenic mutations in other loci. The high penetrance of GJB2 mutations (very much the c.35delG and c.-23+1G>A) in Kurdish population may increase the chance for determining newborns with the ARNSHL by genetic testing. So, it can prevent unnecessary diagnostic testing and improve its diagnosis and treatment in Iran. The Iranian population is composed of various ethnic groups.
Therefore, further studies on the basis of the ethnicity are necessary to identify the ethnicity-based gene profile associated with ARNSHL. So these observations can be helpful in clinical settings to prognosticate the outcomes of genetic testing and disease period to the individuals with HL in Kurdish population.